

Stella Agradi a, *, Majlind Sulce b, Laura Menchetti c, *, Daniele Vigo a, Marta Castrica a, Olimpia Barbato d, Egon Andoni b, Alda Quattrone c, Albana Munga b,
Maria Laura Marongiu e, Giulio Curone a, 1, Gabriele Brecchia a, 1
a Department of Veterinary Medicine and Animal Sciences, University of Milan, Lodi, Italy
b Faculty of Veterinary Medicine, Agricultural University of Tirana, Tirana, Albania
c School of Bioscience and Veterinary Medicine, University of Camerino, Camerino, Italy
d Department of Veterinary Medicine, University of Perugia, Perugia, Italy
e Department of Veterinary Medicine, University of Sassari, Sassari, Italy
a r t i c l e i n f o
Article history:
Received 30 August 2022 Received in revised form 25 March 2023
Accepted 29 March 2023
Available online 25 April 2023
Keywords: Omega 3 Nutraceutical Linseed
Fish oil Algae
a b s t r a c t
Rabbit breeding has many critical aspects related to reproduction, production, and animal welfare, which reduce its profitability as well as consumer attractiveness. Dietary supplementation with n-3 poly- unsaturated fatty acids (PUFA) seems to be a good nutritional strategy to improve several aspects of rabbit breeding, enhance animal welfare and produce a new functional food considered healthy for human consumption. For this reason, the main available scientific research regarding the physiological effects of n-3 PUFA rich products supplemented to the rabbit diet will be reviewed. In particular, con- sequences on the reproductive performances of both doe and buck, the productive parameters, and the meat quality will be analysed.
© 2023 The Authors. Publishing services by Elsevier B.V. on behalf of KeAi Communications Co. Ltd. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-
- Introduction
Rabbit (Oryctolagus cuniculus) breeding has a long history and tradition on the Eurasian continent. To date, China is the world’s largest producer, followed by the Democratic People’s Republic of Korea and the European Union (FAO, 2022). Although rabbit farming has currently slowed in its growth, it still represents a
E-mail addresses: stella.agradi@unimi.it (S. Agradi), laura.menchetti@unicam.it (L. Menchetti).
1 These authors contributed equally. Peer review under responsibility of Chinese Association of Animal Science and Veterinary Medicine.
sector of great economic importance in several geographical areas of the world (FAO, 2022; Wu, 2022). Moreover, the rabbit is nowadays not only intended as a zootechnical species but also as a pet and experimental animal model, widely used in research studies to improve both the productive (Menchetti et al., 2015b, 2019) and reproductive performance of this species (Boiti et al., 1999; Brecchia et al., 2014; Menchetti et al., 2018a), as well to investigate pathophysiology in humans (Collodel et al., 2015; Menchetti et al., 2020a) and other animals (Menchetti et al., 2018b; Zerani et al., 2005). Current research in the cuniculture field is evaluating the influence of farming systems and diets on rabbit ethology, welfare, and meat quality (Menchetti et al., 2020d; Wu, 2022). In particular, during the last decade, the scientific commu- nity has focused on the production of new functional foods and the improvement of reproductive and productive performance by adding different nutraceuticals to the rabbit diet (Menchetti et al., 2019, 2020b). The transformation of rabbit meat into a functional food could be obtained by supplementing rabbit feed with
https://doi.org/10.1016/j.aninu.2023.03.009
2405-6545/© 2023 The Authors. Publishing services by Elsevier B.V. on behalf of KeAi Communications Co. Ltd. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
S. Agradi, M. Sulce, L. Menchetti et al. Animal Nutrition 14 (2023) 70e78
nutraceutical compounds, such as plant-derived products (Agradi et al., 2022; Castrica et al., 2020; Menchetti et al., 2020c), prod- ucts of animal origin (Rodríguez et al., 2017a), and vitamins (Dal Bosco et al., 2004; Lo Fiego et al., 2004). According to the latest research, the compounds that gave the most interesting results are both plant and animal products rich in n-3 polyunsaturated fatty acids (PUFA). Although several biological matrices have been tested, most studies have focused on flax-derived products, fish oil, and algae (Castellini et al., 2019; Curone et al., 2022; Mattioli et al., 2021).
On the other hand, thanks to their physiological multilevel ef- fects (Andoni et al., 2021; Mattioli et al., 2021), adding nutraceut- icals to the rabbit diet can positively influence productive and reproductive performance as well as animal welfare. This aspect is particularly interesting, considering the high mortality and replacement rate of the rabbit does, mainly ascribable to the reproductive rhythms employed in intensive farming (Castellini et al., 2010) which reduce the reproductive and productive per- formance and, thus, the profitability of the farm. Importantly, it is also an indicator of poor animal welfare (Dalmau et al., 2020).
In this review, the main available scientific literature regarding the physiological effects of n-3 PUFA rich products supplemented to the rabbit diet will be discussed, analysing the consequences on the reproductive performance of both doe and buck, the productive parameters, and the meat quality.
- Research methodology
- Literature inclusion criteria
Only peer-reviewed articles or reviews written in English and from WOS and Scopus indexed journals were included.
- Literature identification
The literature search was performed using the most popular research databases in the animal and veterinary sciences field: Web of Science, PubMed, and Google Scholar (consulted until August 2022). The search was conducted utilizing the following keywords: European rabbit, O. cuniculus, rabbit, polyunsaturated fatty acids, n- 3 fatty acids, omega-3 fatty acids, linseed, linseed oil, flax, fish oils, fish liver oils, algae, seaweeds. Initially, the relevance of the study was evaluated considering its title. Subsequently, after a first skimming, the adequacy of the selected articles was determined by evaluating the respective abstracts. This first phase of literature search and identification was performed in parallel by two inde- pendent researchers with expertise in the field. The articles selected by both researchers were moved on to the next phase, while manuscripts deemed valid by one author only were discussed among the researchers to find a common decision. Twenty-nine manuscripts were included in the review at this point. Finally, 5 other manuscripts were identified through backward and forward search. Ultimately, 34 studies were included. In conclusion, these manuscripts were stratified first according to the investigated ef- fects (on reproductive and productive performance or meat qual- ity), and then to the n-3 PUFA source (Fig. 1). Scientific literature on n-3 PUFA effects on reproductive performance was also stratified based on the sex of investigated rabbits.
- The metabolism of n-3 PUFA in mammals
Linoleic (LA, C18:2 n-6) and a-linolenic (ALA, C18:3 n-3) acids are essential fatty acids for vertebrates and are the precursors of
two non-interconvertible PUFA families (n-6 PUFA and n-3 PUFA, respectively), which are obtained through desaturation and lengthening cascade reactions of the carbon chain. The interme- diate and final products of these cascade reactions are the very long chain PUFA (VLC-PUFA), which have important biological roles in the living organism (Burdge, 2018). Two VLC-PUFA, the arachidonic (ARA, C20:4 n-6) and the eicosapentaenoic (EPA, C20:5 n-3) acids, are precursors of eicosanoids such as prosta- glandins and thromboxanes (Saini and Keum, 2018). Docosahex- aenoic (DHA, C22:6 n-3) and docosapentaenoic (DPA, C22:5 n-3) acids are the main constituents of the membrane phospholipids in the retina, brain, reproductive tissues, and gametes (Burdge, 2018). EPA and DHA are considered conditionally essential fatty acids because, even if many vertebrates are physiologically able to synthesize them from ALA, the conversion rate can be so low that dietary integration is needed (Givens, 2009). In the terrestrial ecosystems, only a few organisms, such as lichens and soil algae, are able to synthesize de novo large amounts of n-3 VLC-PUFA (Burdge, 2018). Terrestrial plants, which are at the base of the food chain, are not able to synthesize de novo n-3 VLC-PUFA, but only ALA and AL, which are mainly found in photosynthetic membranes, tissues and seeds such as flaxseed (Burdge, 2018). For this reason, terrestrial herbivores, such as rabbits, must rely on their own conversion to meet the n-3 VLC-PUFA requirements (Pilecky et al., 2021). On the other hand, the main sources of n-3 VLC-PUFA for humans, which are at the top of the food chains, are fish and marine algae, while the requirements for the n-6 VLC- PUFA are met by meat ingestion (Burdge, 2018). Moreover, in humans, a lower n-6 PUFA to n-3 PUFA ratio has been widely proven to be more desirable to decrease the risk of different chronic and inflammatory diseases and mental disorders (Husted and Bouzinova, 2016). However, the n-6/n-3 ratio of ingested PUFA has dramatically increased in recent decades, reaching 20e15:1 ratios in Western countries (Husted and Bouzinova, 2016). In the light of the above, the production of functional food enriched with n-3 could be a new complementary strategy to reduce the incidence of several pathologies and to improve human health.
- The effect of n-3 PUFA on reproductive performance
Cuniculture, is based on the continuous breeding and growth of animals, and, as a result, its success is strictly dependent on the reproductive performance (Rommers et al., 2004). The reproduc- tive success of rabbit does is influenced by the parity order as it is known that nulliparous have higher fertility than both primiparous and multiparous (Martínez-Paredes et al., 2012). The main cause for the reduction of fertility in rabbit does could be attributed to the intensive reproductive rhythms, which induce a negative energy balance (Fortun-Lamothe, 2006; Martínez-Paredes et al., 2012; Menchetti et al., 2015a, 2015b) in addition to genital tract infections (Boiti et al., 1999; Brecchia et al., 2010, 2014; Collodel et al., 2015; Menchetti et al., 2018a). These reproductive issues affect predom- inantly females rather than males, in a percentage of 90% to 10%, respectively. Nevertheless, the rabbit buck’s reproductive perfor- mances are as pivotal as the female’s in order to reach good conception and fertility rates. In the last two decades, different research groups have investigated the supplementation of nutra- ceuticals in the rabbit diet to improve the reproductive perfor- mance of both bucks and does (Andoni et al., 2021; Delgado et al., 2018). Among the different nutraceuticals tested, n-3 PUFA have given interesting results, considering their physiological role in cellular membranes, particularly the ones of the gametes, inflam- matory and immune cells (Burdge, 2018). The following are the most relevant studies regarding the use of flax and its derivatives, fish oil, and algae as a source of n-3 PUFA for the improvement of reproductive performance in rabbit.
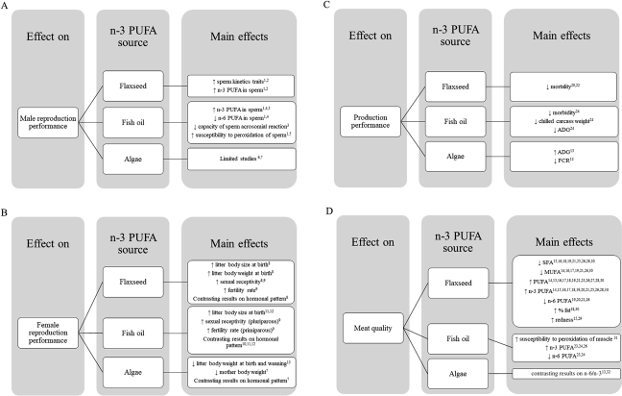
Fig. 1. Main experimental studies investigating n-3 PUFA diet supplementation effects in rabbits.
The studies, which have shown significant effects on the male (A) and female (B) reproductive and productive (C) performance and meat quality (D), are reported. For each aspect, the main effects found in scientific literature have been listed on the base of the n-3 PUFA source (i.e., flaxseed, fish oil or algae) supplemention. The numbers in superscript refer to the experimental studies reporting the specific effect, according to the following legend: 1Castellini et al. (2019); 2Mourvaki et al. (2010); 3Diaz-Fontdevila and Bustos-Obregon (1993); 4Gliozzi et al. (2009); 5Kokoli et al. (2017); 6Kní�zatova/ et al. (2021); 7Okab et al. (2013); 8Elkomy and El-Speiy (2015); 9Mattioli et al. (2019); 10Rebollar et al. (2014); 11Rodríguez et al. (2017b); 12Rodríguez et al. (2018); 13Mordenti et al. (2010); 14Benatmane et al. (2011); 15Bianchi et al. (2006); 16, Dal Bosco et al. (2004); 17Du et al. (2013); 18Tariq et al. (2017); 19Kouba et al. (2008); 20Matics et al. (2017); 21Trebu�sak et al. (2011); 22Casado et al. (2013); 23Rodríguez et al. (2017a); 24Rodríguez et al. (2019); 25Petracci et al. (2009); 26Bernardini et al. (1999); 27Bianchi et al. (2003); 28Bianchi and Cavani (2009); 29Cavani et al.
(2003); 30Peiretti et al. (2010); 31Kowalska and Bielan/ski (2009); 32Peiretti and Meineri (2011). PUFA ¼ polyunsaturated fatty acids; ADG ¼ average daily gain; FCR ¼ feed conversion ratio; SFA ¼ saturated fatty acids; MUFA ¼ monounsaturated fatty acids.
- N-3 PUFA effect on reproductive performance in rabbit buck
VLC-PUFA are synthetised in large amounts in testes and gam- etes of male mammals (among 30% and 50% of the total fatty acids, according to the species) (Burdge, 2018). In particular, the rabbit differs from the majority of other mammals because its sperm has a low presence of DHA, and in turn has a higher level of n-6 DPA, especially with regard to the spermatozoa tail (Mourvaki et al., 2010). Unsaturated phospholipids are of pivotal importance for the spermatozoa plasmatic membrane because they confer fluidity, which assures flexibility to the tail. Indeed, the lipid composition of the membrane regulates the mobility and communication activities of the cell (Castellini et al., 2019). Moreover, the lipid composition of the spermatozoa plasmatic membrane strongly influences the processes of capacitation, acrosome reaction, binding to the zona pellucida, and oocyte fusion (Flesch and Gadella, 2000). The in- clusion of n-3 PUFA in the diet of different species has been shown to affect their sperm characteristics (Lin et al., 2016; Samadian et al., 2010; Van Tran et al., 2017). In these studies, the main aspect to be taken into account, is the quantity of n-3 PUFA supplemented, because higher doses correspond to a higher spermatozoa sus- ceptibility to lipid peroxidation, and lower reproductive perfor- mances (Burdge, 2018). Indeed, sperm has a cytoplasmatic low antioxidant activity. For this reason, it is usually suggested to sup- plement the n-3 PUFA enriched diets with antioxidants such as vitamins E and C. Finally, the period during which the nutraceutical is supplemented should be taken into consideration. A specific amount of time, covering all the spermatogenesis process, is needed to change the spermatozoa plasmatic membrane fatty acid composition. In rabbits, spermatogenesis takes about 50 d from the formation of spermatogonial stem cells to the presence of the same cells in the ejaculated semen (Swierstra and Foote, 1965).
To date, research conducted on the rabbit buck has focused on the evaluation of the effect of a diet enriched with flaxseed, fish oil, and algae, associated or not with antioxidants, on the phospholipid fatty acid profile of the spermatozoa and seminal fluid, semen quality and oxidative status of seminal plasma and sperm. Mourvaki et al. (2010) and Castellini et al. (2019) administered diets supplemented with 5% and 10% of extruded flaxseed, respectively,
to New Zealand White (NZW) male adult rabbits. In both cases, the diet was also enriched with high doses of a-tocopheryl-acetate (200 mg/kg). The vitamin E addition to the diet aims to prevent the
increased susceptibility of spermatozoa to lipid peroxidation as a consequence of the n-3 PUFA supplementation (Castellini et al., 2003). The period of adaptation lasted between 50 and 60 d. Both investigations proved that the flaxseed enriched diet modified the fatty acid profile of sperm, increasing the n-3 PUFA fraction (mainly due to ALA and DHA) and decreasing the n-6 PUFA. The significant increase in DHA testifies the ability of the male gonads to efficiently convert ALA to its VLC-PUFA derivatives. Moreover, the increase in the n-3 PUFA in the plasmatic membrane causes the increment in the membrane fluidity and, thus, the improvement of the motion traits of cells in both studies. Regarding the oxidative stability of the sperm, these studies showed opposite results. The worse oxidative status found in the study by Castellini and colleagues (2019), however, could be attributed to the double percentage of flaxseed supplemented in the diet (10%) compared to the research from Mourvaki et al. (2010) (5%), considering that both diets contained the same over-nutritional amount of vitamin E. Moreover, the study with the higher flaxseed supplementation (10%) also showed an increase in the testosterone blood concentration, ascribable to the phytoestrogens present in the extruded flaxseed, which, acting as hormones, could increase the sex-hormone-binding-globulin syn- thesis, and, as a consequence, the testosterone production (Castellini et al., 2019). On the other hand, Mourvaki et al. (2010) also investigated the modifications in the prostatic granules, whose role is to block sperm capacitation. They found an increase in the number of prostatic granules in the flaxseed rabbit group and hypothesized that this was linked to the increase in the resistance to acrosome reaction of spermatozoa (Mourvaki et al., 2010).
Other studies have investigated the effect of a fish oil enriched diet on the reproductive performance of rabbit buck (Castellini et al., 2003, 2019; Diaz-Fontdevila and Bustos-Obregon, 1993; Gliozzi et al., 2009; Kokoli et al., 2017). The first study was con- ducted in 1993 by Diaz-Fontdevila et al. and investigated the effect of a fish oil supplemented diet on the sperm quality of 4-month-old NZW rabbits divided into 4 equal groups and fed a control diet and 3 experimental diets: (1) a cholesterol-enriched control diet, (2) a cholesterol-PUFA-enriched diet, and (3) a PUFA-enriched diet. PUFA enriched diets consisted of 7.8% (wt:wt) of fish oil containing 22.6% of EPA and 9.7% of DHA. After 60 d of treatment, all the experi- mental diets decreased the kinetics of the acrosomal reaction in live sperm compared to the control. In the case of the only-PUFA- enriched diet group, the authors hypothesized that the elevation in the triglycerides and total lipids in the seminal plasma could explain the reduction in spermatozoa efficiency. However, the high percentage of fish oil inclusion in the diet has to be taken into ac- count. Similar results were obtained by Castellini et al. (2003), which investigated the effect of a diet enriched with 2% fish oil (containing at least 25% of EPA and DPA, and 12.6% of DHA) and/or vitamin E (200 mg/kg) and C (0.5 g/L). The experiment was con- ducted on 30-d-old NZW male rabbits and the diets were admin- istered for 120 d before semen collection. Results showed that the supplemented diet did not affect the volume and concentration of the ejaculate, as well as the characteristics of the fresh spermato- zoa. However, an increase in the susceptibility of semen to perox- idation after storage was shown, which was reduced only when both vitamins E and C were also included in the diet. Modifications in the spermatozoa membrane phospholipids were also demon- strated, with a significant decrease in ARA and an increase in DHA, EPA, and ALA in the rabbits with n-3 PUFA enriched diet. That could probably impair the acrosomal reaction, reducing the fertilization capacity of the semen. These results are consistent with another study by Kokoli et al. (2017) which investigated the effect of a diet enriched with 2% fish oil (containing 9% EPA and 12.5% DHA) for 4 consecutive months on 14 NZW male adult rabbits. Results by Gliozzi et al. (2009) were only partially in agreement with the previously mentioned studies on the fatty acid composition of sperm phospholipids. They involved 5-months-old rabbits fed a diet enriched with 1.5% fish oil (containing 8.3% of EPA, 4.6% of DPA, and 9.4% of DHA) and/or vitamin E (200 mg/kg). The sperm eval- uations were made between 26 to 28 and 37 to 39 weeks of age, thus, after 42 and 119 d from the beginning of the treatment, respectively. The increase of DHA and decrease of LA in the fatty acid composition of sperm were consistent with what was found in the previous study by Castellini et al. (2019). However, the total PUFA and saturated fatty acids decreased and increased respec- tively, contrary to what Castellini et al. (2003) reported. Moreover, controversially, semen susceptibility to lipid peroxidation was not affected by any of the dietary treatments (Gliozzi et al., 2009).
A few studies have evaluated the effect of a selected samples of macroalgae on the reproductive performance of rabbit bucks (Kní�zatova/ et al., 2021; Okab et al., 2013). They showed no patho-
logical signs in both rabbit genders due to the dietary supple- mentation and improvement of some sperm fertility characteristics and prolificacy in does. Despite their encouraging results, these pilot investigations have some limitations, which do not allow to rely on their findings.
Thus, from the analysis of the reported literature, it can be stated that manipulation of the spermatozoa phospholipids fatty acid
profile in rabbit buck can be achieved by diet enrichment with n-3 PUFA for at least an amount of time equal to the spermatogenesis interval. Flaxseed and algae’s use must be further investigated, given the encouraging results so far collected. On the other hand, fish oil seems to cause impaired spermatozoa functionality per se, and to avoid this negative effect, it must be administered with over- nutritional antioxidant levels.
- N-3 PUFA effect on reproductive performance in rabbit doe
Rabbit breeding productivity heavily depends on the repro- ductive performance of the rabbit does. Thus, to maximize the reproduction cycle, the cuniculture breeding systems rely on 3 as- pects: (1) reproduction cycle intensive rhythms, (2) use of artificial insemination, and (3) adoption of more prolific strains (Castellini, 2007). In commercial rabbit production, however, the main cause of culling female rabbits is infertility, which means that the doe refuses service or that there have been 2 to 3 unsuccessful in- seminations (Rosell and de la Fuente, 2009). Indeed, the breeding intensification has led to an alteration of the metabolic and hor- monal status of the female rabbits, due to the overlap of lactation and pregnancy (Fortun-Lamothe, 2006; Martínez-Paredes et al., 2012; Menchetti et al., 2015a, 2015b). This situation is associated with a negative energy balance and hormonal antagonism, which impairs the physiological functioning of the reproductive system (sexual receptivity, ovulation, fertilization, implantation, and em- bryo survival) (Castellini, 2007). Specifically, primiparous does are most affected by this metabolic imbalance because they have to sustain lactation, pregnancy, and body growth at the same time (Parigi Bini and Xiccato, 1998). In this context, n-3 PUFA cover a pivotal role during early pregnancy and the growth of the fetuses, also concerning the visual system and cognitive development. Most lipids are transferred to the fetus via placenta both directly from the diet or through lipolysis (Burdge, 2018). In particular, the rabbit does store n-3 PUFA in different concentrations based on the localization of the adipose tissue. Specifically, the periovarian adi- pose tissue is rich in EPA, DPA and DHA, and could be considered an optimal reservoir of ready-to-use n-3 essential PUFA, available to be employed in physiologic and metabolic ovarian processes (Rebollar et al., 2014). The studies so far conducted on female rabbits fed with enriched n-3 PUFA diets have, indeed, given interesting results, highlighting the important and multilevel involvement of these lipids in every stage of the reproductive cycle.
Regarding the litter-correlated parameters, a first study found higher litter body size and weight at birth in litters from mothers that received the flaxseed-supplemented diet (Elkomy and El- Speiy, 2015). Three mL of flaxseed were administered to does for 7 d previous artificial insemination (Elkomy and El-Speiy, 2015). Three other studies evaluated the use of fish oil as a feed sup- plement (Rebollar et al., 2014; Rodríguez et al., 2017b, 2018). In all these investigations, a commercial n-3 PUFA supplement derived from salmon oil and containing 35% of n-3 PUFA was supple- mented to rabbit does in the amount of 15 g/kg (Rebollar et al., 2014; Rodríguez et al., 2017b) or 60 g/kg (Rodríguez et al., 2018). In the last case, the diet was also supplemented with about 40 mg/kg of vitamin E, instead of 200 mg/kg as in the other 2 investigations. Results from these studies showed no variation (Rebollar et al., 2014) or, in agreement with Elkomy and El-Speiy (2015) even an increase in the kit body sizes (Rodríguez et al., 2017b, 2018). This aspect could have positively affected the new- borns, increasing their survival rate in the first days of life when the localization of nipples is pivotal. However, in contrast with Elkomy and El-Speiy (2015) the litter weight at weaning of sec- ondiparous does that received the n-3 PUFA enriched diet was reduced compared to the control group (Rodríguez et al., 2018).
The authors hypothesized that n-3 PUFA tend to decrease the dimension of abdominal adipose tissue compared to diets rich in MUFA and SFA (Rodríguez et al., 2018). However, an investigation evaluating the effect of both flaxseed (10%) and fish oil (3%) enriched diets on rabbit does, showed a positive effect of both treatments on only the number of weaned rabbits by pluriparous does (Mattioli et al., 2019).
Regarding the effect of seaweed, the first study by Mordenti et al. (2010) evaluated the reproductive and litter performance of nulliparous does given an n-3 PUFA enriched diet with 4 g/kg of marine algae (Schizochytrium spp.) rich in DHA (this amount contributed 750 mg/kg). Regarding the reproductive performance, the only significant results were mean kit weight at birth and at weaning, which decreased in the litters born to mothers that received the supplemented diet. This result could be due to a reduction in the diet palatability caused by the presence of sea- weeds in the feed, which could have negatively affected the reproductive performance of the doe, and, as a consequence, the litter growth. This problem was also evidenced by another study which supplemented the diet of does with 0%, 1% or 2% of the green algae Ulva lactuca (Okab et al., 2013). A decrease in the does mean body weight was shown and ascribed to not only the low digestible nutrients present in the supplemented algae and their high ashes content but also to the reduced palatability of the diet. Thus, regarding the condition of the litters born to mothers that received an n-3 PUFA-supplemented diet, the available scientific literature is not consistent, although feed palatability might be a concern. Moreover, in this context, interesting future research could take into account the fatty acid profile modification in does’ milk. N-3 PUFA enriched diets have usually shown an increase in PUFA and SFA, and specifically in some n-3 PUFA (ALA or DPA and DHA, with flaxseed or fish oil supplementation, respectively) (Mattioli et al., 2019; Rodríguez et al., 2018).
Regarding the other reproductive parameters, the does fertility rate was increased by both flaxseed and fish oil dietary treatment in 2 studies. These studies were performed on pluri- parous does, which usually show poor reproductive performance due to the overlap of lactation and gestation (Mattioli et al., 2019; Rodríguez et al., 2018). Sexual receptivity may also benefit from fish oil and flaxseed supplementation, as shown by Elkomy and El-Speiy (2015) and Mattioli et al. (2019). These aspects could be related to the modification in the hormonal pattern, demon- strated in different studies, even if with different, and sometimes contrasting, results (Elkomy and El-Speiy, 2015; Okab et al., 2013; Rebollar et al., 2014; Rodríguez et al., 2017b, 2018). In this matter, different investigations have evaluated the concentration of blood estrogen in the rabbit doe, starting from the day of insemination until 32 d post-partum (Elkomy and El-Speiy, 2015; Okab et al., 2013; Rodríguez et al., 2017b). All the investigations found an increase in estrogens linked to the administered n-3 PUFA-enriched diet. However, the most interesting hormones to be evaluated are progesterone and prostaglandins. N-3 PUFA are also precursors of the prostaglandins of the 3-series, which have
a lower but similar activity compared to PGF2a. However, at the same time, n-3 PUFA are responsible for inhibiting the synthesis
of eicosanoids deriving from ARA, and, thus, they also reduce the production of PGF2a. PGF2a has a luteolytic activity, and its production can directly influence progesterone synthesis by the
corpus luteum in the ovary. Regarding this topic, the scientific literature is quite inconsistent, because results and speculation have been made in opposite directions (Elkomy and El-Speiy, 2015; Mattioli et al., 2019; Rebollar et al., 2014; Rodríguez et al., 2017b, 2018). Thus, further studies are needed to clarify the effective influence of n-3 PUFA diet integration on the gonadal axis hormonal secretion.
- The effect of n-3 PUFA on productive performance
Productive performance in cuniculture is defined by different parameters related to the health status, growth rate, and metabolic efficiency of the fattening rabbits. In particular, weaning is considered the most critical period for the insurgence of gastroin- testinal diseases which impair the growth rate of animals and require drug use (Curone et al., 2022). However, antimicrobial resistance is becoming a worldwide issue and thus the EU members are incentivizing research on new alternatives to antimicrobials (More, 2020). In this context, the modulation of the intestinal microbiota through diet supplements, including n-3 PUFA, could be an alternative strategy to improve productive performance and the general health status of the animals (Cotozzolo et al., 2020; Cremonesi et al., 2022; Curone et al., 2022; Menchetti et al., 2018b). For these reasons, it is fundamentally important to study new biological compounds that could be used in livestock husbandry to improve their immune system as well as their health and welfare. The supplementation with n-3 PUFA has been investigated as a possible alternative treatment, considering that, thanks to immu- nomodulatory properties, they regulate several homeostatic and inflammatory processes (Castellini et al., 2022; Dennis and Norries, 2015; Mattioli et al., 2021; Menchetti et al., 2020b). Only studies which used isoenergetic diets to evaluate the effect of an n-3 PUFA- enriched feed were considered for reliable results as the utilization
of non-isoenergetic diets can lead to biased results (Trebu�sak et al., 2011).
In general, the studies so far conducted on fattening rabbits have found little influence of n-3 PUFA on the productive performance of the animals, independently whether flaxseed (Benatmane et al., 2011; Bianchi et al., 2006; Dal Bosco et al., 2004; Du et al., 2013;
Eiben et al., 2010; Kouba et al., 2008; Matics et al., 2017; Trebu�sak
et al., 2011), fish oil (Rodríguez et al., 2017a, 2019) or seaweeds are used (Dalle Zotte et al., 2013).
The first study conducted utilizing flaxseed as an n-3 PUFA
source was by Dal Bosco et al. (2004). This investigation evaluated the effect of an 8% flaxseed enriched diet (with 200 mg/kg of a- tocopheryl acetate) on the productive performance of fattening
rabbits from 45 d of life until slaughter (at 85 d). No differences were found with respect to the control group. The same was shown in other studies evaluating the effect of flaxseed at different con- centrations between 3% and 9% on final body weight, average daily gain, feed intake, and feed conversion ratio (Benatmane et al., 2011;
Bernardini et al., 1999; Bianchi et al., 2006; Eiben et al., 2010; Kouba et al., 2008; Matics et al., 2017; Trebu�sak et al., 2011, 2019). How- ever, a decrease in mortality between 5 and 11 weeks was shown in
the study by Matics et al. (2017) which evaluated the effect of a 1 to 4 weeks duration diet enriched with 3% flaxseed oil. This result confirmed the study by Casado et al. (2013).
With regard to the use of fish oil in the fattening rabbit diet, two studies by Rodríguez et al. (2017a, 2019) have investigated the ef- fect of salmon oil derivate rich in EPA and DHA, supplemented at 1.5% in the diet of rabbits weaned at 25 or 30 d of age. The diet was administered to rabbit does and their litters until slaughtering. In the first study, no effect was found on the productive performances (Rodríguez et al., 2017a), whereas in the second one, a decrease in the average daily gain, as well as hot and chilled carcass weight, was shown (Rodríguez et al., 2019). However, the morbidity was found significantly lower in the fish oil-supplemented group compared to the control one, and this result could be linked to the inflammation regulation actuated by n-3 PUFA and their metabolites (Rodríguez et al., 2019).
Finally, some investigations have assessed the effect of seaweed inclusion in the fattening rabbit diet. The study by Dalle Zotte et al. (2013) showed no effect of Spirulina alga (Arthrospira platensis) on
the productive performance of companion rabbits. However, this investigation was conducted on pet animals, and the results about productive performance could be biased because these rabbit breeds are not selected for fattening. A second study evaluated the supplementation of 4 g/kg of marine algae (Schizochytrium spp.) rich in DHA (this amount contributed 750 mg/kg of DHA) to nulliparous does and their litters until slaughtering (Mordenti et al., 2010). This study showed positive effects, with an increase in the average daily gain and a decrease in the feed conversion ratio in fattening rabbits born to mothers which received the enriched diet (independently from the diet they were fed during growing and finishing periods). These results suggested that the addition of n-3 PUFA in the rabbit does diet could modify their milk composition and strengthen the litter’s defence mechanisms. Indeed, manipu- lation of the fatty acid profile of rabbit does’ milk via diet has been shown in different studies (Mattioli et al., 2019; Rodríguez et al., 2018).
Thus, in summary, it seems that n-3 PUFA supplemented diet administered to fattening rabbits did not affect their productive performance, independently from the source. However, some interesting effects have been shown on the mortality and/or morbidity of growing rabbits, and that aspect should be further investigated along with the influence of n-3 PUFA on the milk, immune status and gut microbiota, pivotal pillars during the critical weaning period.
- The effect of n-3 PUFA on meat quality
Rabbit meat is extremely lean and low in calories. It has an excellent percentage of high biological value proteins and at the same time a very low quantity of cholesterol. SFAs are the pre- dominant fatty acids, although rabbit meat contains high amounts of MUFA and PUFA, compared to other livestock meats (Poławska et al., 2013). For these characteristics, rabbit meat is highly appre- ciated by consumers, at least in some geographical areas (i.e. Europe, Mediterranean Africa, Asia, and some Latin American countries) (McNitt et al., 2013). However, to date, the rabbit sector is experiencing a hard economic crisis due to structural weak- nesses, progressive and constant reductions in consumption, and rising criticism by Western consumers related to animal welfare conditions and other ethical issues. Moreover, the price of rabbit meat is still higher compared to other kinds of meat such as poultry or pork (Hoffman et al., 2004). A very important factor for some consumers in choosing meat is the welfare of the animals raised. In this context, the use of rabbit meat as a functional food could be important to relaunch the rabbit sector and the supplementation of n-3 PUFA is a strategy widely tested to produce functional food. Importantly, these molecules could also improve the rabbit’s health and welfare. Monogastric animals have proved to be the most successful in the production of functional food, thanks to their direct absorption of lipids supplemented in the diet without alteration in the digestive system. However, the main issue faced by researchers in enriching animal-derived products with n-3 PUFA is the increase of their oxidative status (O’Keefe et al., 1995). Inde- pendently from the species involved, this problem has been shown to negatively affect the shelf-life of the final product and the pur- chase by the consumer, because of the off-flavors originating from lipid peroxidation, and further technologic processing (Ahn et al., 1996). For these reasons, many studies have investigated the oxidative status of the products along with the fatty acid profile
modifications in animal tissues (Benatmane et al., 2011; Dal Bosco et al., 2004; Kowalska and Bielan/ski, 2009). To avoid or reduce these issues, many authors have added antioxidants, such as
vitamin E (Dal Bosco et al., 2004). Finally, it is important to deter- mine the amount of time needed to modify, via an enriched diet,
the fatty acid profile of the products. For flaxseed supplementation in the rabbit, it has been established at 2 to 3 weeks before slaughtering (Matics et al., 2017).
To date, in the cuniculture research field, many investigations have been conducted to evaluate the possible production of a functional food through the administration of n-3 PUFA-enriched diets to fattening rabbits. The great majority of the investigations used flaxseed as a source of ALA, with a supplementation between 1.6% and 16% (Benatmane et al., 2011; Bernardini et al., 1999; Bianchi et al., 2003, 2006; Bianchi and Cavani, 2009; Cavani et al., 2003; Dal Bosco et al., 2004; Du et al., 2013; Eiben et al., 2010; Kouba et al., 2008; Matics et al., 2017; Peiretti et al., 2010; Petracci
et al., 2009; Tariq et al., 2017; Trebu�sak et al., 2011). A few studies
evaluated the effect of fish oil on rabbit meat quality (Bernardini et al., 1999; Kowalska and Bielan/ski, 2009; Rodríguez et al., 2017a, 2019), and, to our knowledge, just 2 studies used seaweed as n-3
PUFA sources (Mordenti et al., 2010; Peiretti and Meineri, 2011). In general, the results about the physico-chemical characteris-
tics of the rabbit meat (usually evaluated on the loin meat) showed a few effects. Only a few authors have found an increase in the meat redness due to the flaxseed-supplemented diet (at 8%) (Bianchi et al., 2006; Cavani et al., 2003). The chemical composition of the supplemented rabbit meat was in general not affected by the n-3 PUFA diet inclusion, even if some studies reported an increase in the lipid percentage (Peiretti and Meineri, 2011; Tariq et al., 2017). The results are more robust as regards the meat fatty acid pro- file. In several studies the supplementation of rabbit diet with n-3 PUFA has shown to be effective in enriching meat with both n-3 PUFA and n-3 VLC-PUFA, testifying the ability of rabbit tissues to desaturate and elongate the precursor (ALA) of n-3 VLC-PUFA into these latter molecules (Benatmane et al., 2011; Bernardini et al., 1999; Bianchi et al., 2006; Dal Bosco et al., 2004; Du et al., 2013;
Kouba et al., 2008; Matics et al., 2017; Tariq et al., 2017; Trebu�sak
et al., 2011). For this reason, a diet enriched with flaxseed is able to increase mainly the meat concentration of PUFA, n-3 PUFA, with particular reference to ALA, and decrease the one of SFA and MUFA (indeed n-3 PUFA have been proven to exert an inhibitory action on stearoyl-CoA-desaturase) (Kouba et al., 2008). In enriched n-3 PUFA meat, the n-3 VLC-PUFA, such as EPA, DPA and DHA, have also shown increased values compared to the control groups, even if they always resulted to be present in very low concentrations (Benatmane et al., 2011; Bernardini et al., 1999; Bianchi et al., 2006; Dal Bosco et al., 2004; Du et al., 2013; Kouba et al., 2008; Matics
et al., 2017; Tariq et al., 2017; Trebu�sak et al., 2011). A decrease of
the n-6/n-3 ratio has also been achieved, with values indirectly proportional to the supplemented quantity of flaxseed, reaching values of 1.5 with feed supplementation higher than 8%, which are
considered very good for human health (Peiretti et al., 2010; Trebu�sak et al., 2011). Moreover, the atherogenicity and thrombo- genicity indexes have shown lower and healthier values for the
meat of rabbits that received the flaxseed enriched diet (Dal Bosco et al., 2004; Peiretti et al., 2010; Rodríguez et al., 2017a). However, it has to be considered that the fatty acid profile of meat is also affected by the muscle analysed, from the percentage of n-3 PUFA inclusion in the diet and its duration, and from the process which the meat undergoes after slaughtering (Matics et al., 2017).
The few experiments which assessed the effect on meat fatty acid profile of a fish oil-enriched diet in rabbits have shown a common increase in PUFA, n-3 PUFA, and n-3 VLC-PUFA, with the n-6/n-3 ratio reaching 1.61 (Bernardini et al., 1999; Kowalska and
Bielan/ski, 2009; Rodríguez et al., 2017a, 2019).
Regarding the utilization of seaweed, the scarce scientific liter- ature has shown contrasting results depending on the algae sup- plemented to the diet (Mordenti et al., 2010; Peiretti and Meineri, 2011). Indeed, even if this ingredient can bring high amounts of
n-3 VLC-PUFA, it could be also responsible for the increase in n-6 PUFA. This could be caused by some species of algae that contain a high concentration of LA, which increase n-6/n-3 ratio rather than decreasing it (Peiretti and Meineri, 2011).
However, as previously stated, when enriching a food product with PUFA, it is essential to consider the possible increased risk for lipid peroxidation, which could affect not only the health of the animal but also many aspects related to the final food product. In the majority of the studies conducted to date, lipid susceptibility to oxidation has been shown to increase in the meat of rabbits which received the n-3 PUFA-enriched diet (Benatmane et al., 2011; Bianchi et al., 2003, 2006; Bianchi and Cavani, 2009; Cavani et al.,
2003; Kowalska and Bielan/ski, 2009; Trebu�sak et al., 2011).
Bianchi et al. (2006, 2003), also based on a sensory test, suggested that the shelf-life limit for industrial storage of n-3 PUFA enriched meat is 6 months. Different investigations have tried to prevent the lipid oxidation phenomena by supplementing the feed also with vitamin E, which is a natural antioxidant. It has been proved that 30 mg/kg of vitamin E in the feed is insufficient to limit the dete- rioration of the meat, while 100 and 200 mg/kg have been demonstrated to be effective in the preservation of the product
after 90 d of frozen storage or 8 d of fresh storage, respectively (Bianchi et al., 2003; Kowalska and Bielan/ski, 2009).
In conclusion, flaxseed supplementation for 2 to 3 weeks before slaughtering, when accompanied by vitamin E addition, seems to be the most effective, cheap, and sustainable way to obtain an n-3 PUFA enriched rabbit meat, which could be a useful tool to improve the n-6/n-3 ratio in the human diet. However, further research should be conducted to establish the correct amount of flaxseed and vitamin E to be supplemented to obtain a good product both from a nutritional and a technological point of view.
- Conclusions and perspectives
To date, the available scientific literature on the use of n-3 PUFA in rabbit diets has demonstrated the effectiveness of this nutri- tional strategy to improve several aspects related not only to male and female reproductive traits (such as spermatozoa structure, pluriparous does fertility, sexual receptivity, mortality and morbidity of litters) but also to the fatty acid profile of rabbit meat, specifically the n-6/n-3 ratio and percentage of essential fatty acids. Moreover, particularly interesting are also the aspects related to the modification of the intestinal microbial flora which in turn induces stimulation of the immune system which reduces the risk of dis- ease and consequently confers greater profits for the farmer, better welfare for the animals, and reduced risks for the consumer related to antibiotic resistance and residues in meat. The n-3 PUFA source that has given the most interesting results, considering its com- mercial and economic availability, is flaxseed. Nevertheless, a concomitant supplementation with some antioxidants, such as vitamin E should always be taken into account.
Further research will be needed to better understand the physiological mechanisms which underlie both the improvement of some aspects and the prevention of the oxidative instability deriving from n-3 PUFA-enriched animal tissues. In conclusion, general results suggest that the use of n-3 PUFA and vitamin E in cuniculture should be incentivized as a tool to relaunch the entire sector.
Author contributions
Stella Agradi, Laura Menchetti, Giulio Curone, Gabriele Brecchia designed the research. Stella Agradi, Majlind Sulce, Daniele Vigo, Marta Castrica, Egon Andoni, Gabriele Brecchia performed the research. Laura Menchetti, Olimpia Barbato,
Albana Munga, Maria Laura Marongiu analyzed the data. Stella Agradi, Laura Menchetti, Alda Quattrone, Maria Laura Marongiu, Giulio Curone, Gabriele Brecchia wrote the paper. Stella Agradi, Gabriele Brecchia had primary responsibility for the final content. All authors read and approved the final manuscript.
Declaration of competing interest
We declare that we have no financial and personal relationships with other people or organizations that can inappropriately influ- ence our work, and there is no professional or other personal in- terest of any nature or kind in any product, service and/or company that could be construed as influencing the content of this paper.
Acknowledgement
This research was supported by Fondo di Ateneo per la Ricerca 2019 FAR2019MARONGIU e Studio di sistemi alternativi di gestione della fase riproduttiva rispettosi della fisiologia e del benessere della coniglia- of University of Sassari.
References
Agradi S, Draghi S, Cotozzolo E, Barbato O, Castrica M, Quattrone A, et al. Goji berries supplementation in the diet of rabbits and other livestock animals: a mini-review of the current knowledge. Front Vet Sci 2022;8:823589. https:// doi.org/10.3389/fvets.2021.823589.
Ahn DU, Lutz S, Sim JS. Effects of dietary a-linolenic acid on the fatty acid composition, storage stability and sensory characteristics of pork loin. Meat Sci
1996;43:291e9. https://doi.org/10.1016/S0309-1740(96)00001-0.
Andoni E, Curone G, Agradi S, Barbato O, Menchetti L, Vigo D, et al. Effect of goji berry (lycium barbarum) supplementation on reproductive performance of rabbit does. Animals 2021;11:1672. https://doi.org/10.3390/ani11061672.
Benatmane F, Kouba M, Youyou A, Mourot J. Effect of a linseed diet on lipogenesis, fatty acid composition and stearoyl-CoA-desaturase in rabbits. Animal 2011;5: 1993e2000. https://doi.org/10.1017/S1751731111001145.
Bernardini M, Dal Bosco A, Castellini C. Effect of dietary n-3/n-6 ratio on fatty acid composition of liver, meat and perirenal fat in rabbits. Anim Sci 1999;68: 647e54. https://doi.org/10.1017/S1357729800050682.
Bianchi M, Cavani C. The influence of linseed on rabbit meat quality. World Rabbit Sci 2009;17:97e107. https://doi.org/10.4995/wrs.2009.663.
Bianchi M, Betti M, Petracci M, Cavani C. Technological characteristics of rabbit meat with raised n-3 PUFA Technological characteristics of rabbit meat with raised n- 3 PUFA. Ital J Anim Sci 2003;2:444e6. https://doi.org/10.4081/ijas.2003.s1.444.
Bianchi M, Petracci M, Cavani C. Effects of dietary inclusion of dehydrated lucerne and whole linseed on rabbit meat quality. World Rabbit Sci 2006;14:247e58. https://doi.org/10.4995/wrs.2006.562.
Boiti C, Canali C, Brecchia G, Zanon F, Facchin E. Effects of induced endometritis on the life-span of corpora lutea in pseudopregnant rabbits and incidence of spontaneous uterine infections related to fertility of breeding does. Ther- iogenology 1999;52:1123e32. https://doi.org/10.1016/S0093-691X(99)00204-6. Brecchia G, Cardinali R, Mourvaki E, Collodel G, Moretti E, Dal Bosco A, et al. Short- and long-term effects of lipopolysaccharide-induced inflammation on rabbit sperm quality. Anim Reprod Sci 2010;118:310e6. https://doi.org/10.1016/
Brecchia G, Menchetti L, Cardinali R, Castellini C, Polisca A, Zerani M, et al. Effects of a bacterial lipopolysaccharide on the reproductive functions of rabbit does. Anim Reprod Sci 2014;147:128e34. https://doi.org/10.1016/j.anireprosci.2014. 04.009.
Burdge GC. Polyunsaturated fatty acid metabolism. 1st ed. Academic Press and AOCS Press; 2018. https://doi.org/10.1016/C2016-0-00749-0.
Casado C, Moya VJ, Pascual JJ, Blas E, Cervera C. Dietary fatty acid profile: effects on caecal fermentation and performance of young and fattening rabbits. World Rabbit Sci 2013;21:235e42. https://doi.org/10.4995/wrs.2013.1437.
Castellini C. Reproductive activity and welfare of rabbit does. Ital J Anim Sci 2007;6: 743e7. https://doi.org/10.4081/ijas.2007.1s.743.
Castellini C, Lattaioli P, Dal Bosco A, Minelli A, Mugnai C. Oxidative status and semen characteristics of rabbit buck as affected by dietary vitamin E, C and n-3 fatty acids. Reprod, Nutr, Dev 2003;43:91e103. https://doi.org/10.1051/rnd:2003008.
Castellini C, Dal Bosco A, Arias-A/lvarez M, Lorenzo PL, Cardinali R, Rebollar PG. The
main factors affecting the reproductive performance of rabbit does: a review. Anim Reprod Sci 2010;122:174e82. https://doi.org/10.1016/j.anireprosci.2010. 10.003.
Castellini C, Mattioli S, Signorini C, Cotozzolo E, Noto D, Moretti E, et al. Effect of dietary n-3 source on rabbit male reproduction. Oxid Med Cell Longev 2019;2019:3279670. https://doi.org/10.1155/2019/3279670.
Castellini C, Mattioli S, Moretti E, Cotozzolo E, Perini F, Dal Bosco A, et al. Expression of genes and localization of enzymes involved in polyunsaturated fatty acid
synthesis in rabbit testis and epididymis. Sci Rep 2022;12:2637. https://doi.org/ 10.1038/s41598-022-06700-y.
Castrica M, Menchetti L, Balzaretti CM, Branciari R, Ranucci D, Cotozzolo E, et al. Impact of dietary supplementation with goji berries (lycium barbarum) on microbiological quality, physico-chemical, and sensory characteristics of rabbit meat. Foods 2020;9:1480. https://doi.org/10.3390/foods9101480.
Cavani C, Betti M, Bianchi M, Petracci M. Effects of the dietary inclusion of vegetable fat and dehydrated alfalfa meal on the technological properties of rabbit meat. Vet Res 2003;27:643e6.
Collodel G, Moretti E, Brecchia G, Ku�zelov/a L, Arruda J, Mourvaki E, et al. Cytokines
release and oxidative status in semen samples from rabbits treated with bac- terial lipopolysaccharide. Theriogenology 2015;83:1233e40. https://doi.org/ 10.1016/j.theriogenology.2015.01.008.
Cotozzolo E, Cremonesi P, Curone G, Menchetti L, Riva F, Biscarini F, et al. Charac- terization of bacterial microbiota composition along the gastrointestinal tract in rabbits. Animals 2020;11:31. https://doi.org/10.3390/ani11010031.
Cremonesi P, Curone G, Biscarini F, Cotozzolo E, Menchetti L, Riva F, et al. Dietary supplementation with goji berries (lycium barbarum) modulates the microbiota of digestive tract and caecal metabolites in rabbits. Animals 2022;12:121.
Curone G, Biscarini F, Cotozzolo E, Menchetti L, Dal Bosco A, Riva F, et al. Could dietary supplementation with different sources of N-3 polyunsaturated fatty acids modify the rabbit gut microbiota? Antibiotics 2022;11:1e21. https:// doi.org/10.3390/antibiotics11020227.
Dal Bosco A, Castellini C, Bianchi L, Mugnai C. Effect of dietary a-linolenic acid and vitamin E on the fatty acid composition, storage stability and sensory traits of
rabbit meat. Meat Sci 2004;66:407e13. https://doi.org/10.1016/S0309-1740(03) 00127-X.
Dalmau A, Moles X, Pallisera J. Animal welfare assessment protocol for does, bucks, and kit rabbits reared for production. Front Vet Sci 2020;7:445. https://doi.org/ 10.3389/fvets.2020.00445.
Dalle Zotte A, Sartori A, Bohatir P, Re/mignon H, Ricci R. Effect of dietary supple-
mentation of Spirulina (Arthrospira platensis) and Thyme (Thymus vulgaris) on growth performance, apparent digestibility and health status of companion dwarf rabbits. Livest Sci 2013;152:182e91. https://doi.org/10.1016/ j.livsci.2012.12.017.
Delgado R, Abad-Guama/n R, Nicodemus N, Villamide MJ, Ruiz-Lo/pez N, Caraban~o R,
et al. Effect of level of soluble fiber and n-6/n-3 fatty acid ratio on performance of rabbit does and their litters. J Anim Sci 2018;96:1084e100. https://doi.org/ 10.1093/jas/sky019.
Dennis EA, Norries PC. Eicosanoid storm in infection and inflammation. Nat Rev Immunol 2015;15:511e23. https://doi.org/10.1038/nri3859.
Diaz-Fontdevila M, Bustos-Obregon E. Cholesterol and polyunsaturated acid enriched diet: effect on kinetics of the acrosome reaction in rabbit spermatozoa. Mol Reprod Dev 1993;35:176e80. https://doi.org/10.1002/mrd.1080350211.
Du HT, Wang CY, Wang XP, Ma MW, Li FC. The effects of dietary a-linolenic acid on growth performance, meat quality, fatty acid composition and liver relative
enzyme mRNA expression of growing meat rabbits. J Anim Feed Sci 2013;22: 122e9. https://doi.org/10.22358/jafs/66002/2013.
Eiben C, Ve/gi B, Vira/g G, Go/dor-Surmann K, Maro/ A, Odermatt M, et al. Effect of
different dietary ratios of sunflower and linseed oils on growth and carcass traits of rabbits. Livest Sci 2010;131:15e22. https://doi.org/10.1016/j.livsci.2010. 02.017.
Elkomy AE, El-Speiy ME. Polyunsaturated fatty acids combined with equine cho- rionic gonadotropin to enhance reproductive performance in aged rabbit does. Ital J Anim Sci 2015;14:39e44. https://doi.org/10.4081/ijas.2015.3535.
FAO. FAOSTAT. FAO Dep Off 2022, https://www.fao.org/faostat/en/#data/QCL.
Lo Fiego DP, Santoro P, Macchioni P, Mazzoni D, Piattoni F, Tassone F, et al. The effect of dietary supplementation of vitamins C and E on the a-tocopherol content of muscles, liver and kidney, on the stability of lipids, and on certain meat quality
parameters of the longissimus dorsi of rabbits. Meat Sci 2004;67:319e27. https://doi.org/10.1016/j.meatsci.2003.11.004.
Flesch FM, Gadella BM. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. Biochim Biophys Acta Rev Biomembr 2000;1469: 197e235. https://doi.org/10.1016/S0304-4157(00)00018-6.
Fortun-Lamothe L. Energy balance and reproductive performance in rabbit does. Anim Reprod Sci 2006;93:1e15. https://doi.org/10.1016/j.anireprosci.2005.06. 009.
Givens I. Animal nutrition and lipids in animal products and their contribution to human intake and health. Nutrients 2009;1:71e82. https://doi.org/10.3390/ nu1010071.
Gliozzi TM, Zaniboni L, Maldjian A, Luzi F, Maertens L, Cerolini S. Quality and lipid composition of spermatozoa in rabbits fed DHA and vitamin E rich diets. Theriogenology 2009;71:910e9. https://doi.org/10.1016/j.theriogenology.2008. 10.022.
Hoffman L, Nkhabutlane P, Schutte D, Vosloo C. Factors affecting the purchasing of rabbit meat: a study of ethnic groups in the Western Cape. J Fam Ecol Consum Sci 2004;32:26e35. https://doi.org/10.4314/jfecs.v32i1.52850.
Husted KS, Bouzinova EV. The importance of n-6/n-3 fatty acids ratio in the major depressive disorder. Med 2016;52:139e47. https://doi.org/10.1016/j.medici. 2016.05.003.
Kní�zatov/a N, Massa/nyi M, Tok/arov/a K, Vizzarri F, Massa/nyi P, Ondru�ska L�, et al.
Effect of dietary supplementation with seaweed and polyphenols mixture on antioxidant status, concentration and motility of rabbit spermatozoa. J Microbiol Biotechnol Food Sci 2021;10:1e4. https://doi.org/10.15414/ jmbfs.2179.
Kokoli AN, Lavrentiadou SN, Zervos IA, Tsantarliotou MP, Georgiadis MP, Nikolaidis EA, et al. Dietary omega-3 polyunsaturated fatty acids induce plas- minogen activator activity and DNA damage in rabbit spermatozoa. Andrologia 2017;49:1e9. https://doi.org/10.1111/and.12776.
Kouba M, Benatmane F, Blochet JE, Mourot J. Effect of a linseed diet on lipid oxidation, fatty acid composition of muscle, perirenal fat, and raw and cooked rabbit meat. Meat Sci 2008;80:829e34. https://doi.org/10.1016/j.meatsci.2008. 03.029.
Kowalska D, Bielan/ski P. Meat quality of rabbits fed a diet supplemented with fish
oil and antioxidant Meat quality of rabbits fed a diet supplemented with fish oil and antioxidant. Anim Sci Pap Rep 2009;27:139e48.
Lin Y, Cheng X, Mao J, Wu D, Ren B, Xu SY, et al. Effects of different dietary n-6/n-3 polyunsaturated fatty acid ratios on boar reproduction. Lipids Health Dis 2016;15:1e10. https://doi.org/10.1186/s12944-016-0193-8.
Martínez-Paredes E, Ro/denas L, Martínez-Vallespín B, Cervera C, Blas E, Brecchia G,
et al. Effects of feeding programme on the performance and energy balance of nulliparous rabbit does. Animal 2012;6:1086e95. https://doi.org/10.1017/ S1751731111002643.
Matics Z, Cullere M, Szín M, Gerencse/r Z, Szabo/ A, Fe/bel H, et al. Effect of a dietary
supplementation with linseed oil and selenium to growing rabbits on their productive performances, carcass traits and fresh and cooked meat quality. J Anim Physiol Anim Nutr 2017;101:685e93. https://doi.org/10.1111/jpn.12589.
Mattioli S, Dal Bosco A, Maranesi M, Petrucci L, Rebollar PG, Castellini C. Dietary fish oil and flaxseed for rabbit does: fatty acids distribution and D6-desaturase enzyme expression of different tissues. Animal 2019;13:1934e42. https://
doi.org/10.1017/S175173111900020X.
Mattioli S, Collodel G, Signorini C, Cotozzolo E, Noto D, Cerretani D, et al. Tissue antioxidant status and lipid peroxidation are related to dietary intake of n-3 polyunsaturated acids: a rabbit model. Antioxidants 2021;10:681. https:// doi.org/10.3390/antiox10050681.
McNitt JI, Lukefahr SD, Cheeke PR, Patton NM. Rabbit production. 9th ed. Oxford- shire: CABI Publishers; 2013. https://doi.org/10.4995/wrs.2013.1674.
Menchetti L, Brecchia G, Canali C, Cardinali R, Polisca A, Zerani M, et al. Food re- striction during pregnancy in rabbits: effects on hormones and metabolites involved in energy homeostasis and metabolic programming. Res Vet Sci 2015a;98:7e12. https://doi.org/10.1016/j.rvsc.2014.11.017.
Menchetti L, Brecchia G, Cardinali R, Polisca A, Boiti C. Feed restriction during pregnancy: effects on body condition and productive performance of primip- arous rabbit does. World Rabbit Sci 2015b;23:1e8. https://doi.org/10.4995/ wrs.2015.1703.
Menchetti L, Barbato O, Filipescu IE, Traina G, Leonardi L, Polisca A, et al. Effects of local lipopolysaccharide administration on the expression of Toll-like receptor 4 and pro-inflammatory cytokines in uterus and oviduct of rabbit does. Ther- iogenology 2018a;107:162e74. https://doi.org/10.1016/j.theriogenology.2017.10. 046.
Menchetti L, Canali C, Castellini C, Boiti C, Brecchia G. The different effects of linseed and fish oil supplemented diets on insulin sensitivity of rabbit does during pregnancy. Res Vet Sci 2018b;118:126e33. https://doi.org/10.1016/j.rvsc.2018. 01.024.
Menchetti L, Vecchione L, Filipescu I, Petrescu VF, Fioretti B, Beccari T, et al. Effects of Goji berries supplementation on the productive performance of rabbit. Livest Sci 2019;220:123e8. https://doi.org/10.1016/j.livsci.2018.12.016.
Menchetti L, Andoni E, Barbato O, Canali C, Quattrone A, Vigo D, et al. Energy ho- meostasis in rabbit does during pregnancy and pseudopregnancy. Anim Reprod Sci 2020a;218:106505. https://doi.org/10.1016/j.anireprosci.2020.106505.
Menchetti L, Barbato O, Sforna M, Vigo D, Mattioli S, Curone G, et al. Effects of diets enriched in linseed and fish oil on the expression pattern of toll-like receptors 4 and proinflammatory cytokines on gonadal Axis and reproductive organs in rabbit buck. Oxid Med Cell Longev 2020b;2020:4327470. https://doi.org/ 10.1155/2020/4327470.
Menchetti L, Brecchia G, Branciari R, Barbato O, Fioretti B, Codini M, et al. The effect of Goji berries (Lycium barbarum) dietary supplementation on rabbit meat quality. Meat Sci 2020c;161:108018. https://doi.org/10.1016/j.meatsci.2019. 108018.
Menchetti L, Curone G, Andoni E, Barbato O, Troisi A, Fioretti B, et al. Impact of goji berries (Lycium barbarum) supplementation on the energy homeostasis of rabbit does: uni- and multivariate approach. Animals 2020d;10:1e19. https:// doi.org/10.3390/ani10112000.
Mordenti AL, Sardi L, Bonaldo A, Pizzamiglio V, Brogna N, Cipollini I, et al. Influence of marine algae (Schizochytrium spp.) dietary supplementation on doe per- formance and progeny meat quality. Livest Sci 2010;128:179e84. https:// doi.org/10.1016/j.livsci.2009.12.003.
More SJ. European perspectives on efforts to reduce antimicrobial usage in food animal production. Ir Vet J 2020;73:1e12. https://doi.org/10.1186/s13620-019- 0154-4.
Mourvaki E, Cardinali R, Dal Bosco A, Corazzi L, Castellini C. Effects of flaxseed di- etary supplementation on sperm quality and on lipid composition of sperm subfractions and prostatic granules in rabbit. Theriogenology 2010;73:629e37. https://doi.org/10.1016/j.theriogenology.2009.10.019.
Okab AB, Samara EM, Abdoun KA, Rafay J, Ondruska L, Parkanyi V, et al. Effects of dietary seaweed (Ulva lactuca) supplementation on the reproductive perfor- mance of buck and doe rabbits. J Appl Anim Res 2013;41:347e55. https:// doi.org/10.1080/09712119.2013.783479.
O’Keefe SF, Proudfoot FG, Ackman RG. Lipid oxidation in meats of omega-3 fatty acid-enriched broiler chickens. Food Res Int 1995;28:417e24. https://doi.org/ 10.1016/0963-9969(95)00021-D.
Parigi Bini R, Xiccato G. Energy metabolism and requirements. In: De Blas C, Wiseman J, editors. Nutr. Rabbit. Wallingford Oxon, UK: CABI Publishing; 1998.
Peiretti PG, Meineri G. Effects of diets with increasing levels of Spirulina platensis on the carcass characteristics , meat quality and fatty acid composition of growing rabbits. Livest Sci 2011;140:218e24. https://doi.org/10.1016/ j.livsci.2011.03.031.
Peiretti PG, Meineri G, Nazionale C, Animali P, Ecologia E. Effects of diets with increasing levels of golden flaxseed on carcass characteristics , meat quality and lipid traits of growing rabbits. Ital J Anim Sci 2010;9:e70. https://doi.org/ 10.4081/ijas.2010.e70.
Petracci M, Bianchi M, Cavani C. Development of rabbit meat products fortified with n-3 polyunsaturated fatty acids. Nutrients 2009;1:111e8. https://doi.org/ 10.3390/nu1020111.
Pilecky M, Z/avorka L, Arts MT, Kainz MJ. Omega-3 PUFA profoundly affect neural,
physiological, and behavioural competences e implications for systemic changes in trophic interactions. Biol Rev 2021;96:2127e45. https://doi.org/ 10.1111/brv.12747.
Poławska E, Cooper RG, Jo//zwik A, Pomianowski J. Meat from alternative species –
nutritive and dietetic value, and its benefit for human health – a review. CyTA-J Food 2013;11:37e42. https://doi.org/10.1080/19476337.2012.680916.
Rebollar PG, García-García RM, Arias-A/lvarez M, Milla/n P, Rey AI, Rodríguez M, et al.
Reproductive long-term effects, endocrine response and fatty acid profile of rabbit does fed diets supplemented with n-3 fatty acids. Anim Reprod Sci 2014;146:202e9. https://doi.org/10.1016/j.anireprosci.2014.02.021.
Rodríguez M, Carro MD, Valiente V, Formoso-Rafferty N, Rebollar PG. Effects of dietary fish oil supplementation on performance, meat quality, and cecal fermentation of growing rabbits. J Anim Sci 2017a;95:3620e30. https://doi.org/ 10.2527/jas.2017.1690.
Rodríguez M, García-García RM, Arias-A/lvarez M, Formoso-Rafferty N, Mill/an P,
Lo/pez-Tello J, et al. A diet supplemented with -3 polyunsaturated fatty acids influences the metabomscic and endocrine response of rabbit does and their offspring. J Anim Sci 2017b;95:2690. https://doi.org/10.2527/jas2017.1429.
Rodríguez M, García-García RM, Arias-A/lvarez M, Milla/n P, Febrel N, Formoso-
Rafferty N, et al. Improvements in the conception rate, milk composition and embryo quality of rabbit does after dietary enrichment with n-3 poly- unsaturated fatty acids. Animal 2018;12:2080e8. https://doi.org/10.1017/ S1751731117003706.
Rodríguez M, Carro MD, Valiente V, Formoso-Rafferty N, Rebollar PG. Supplemen- tation with fish oil improves meat fatty acid profile although impairs growth performance of early weaned rabbits. Animals 2019;9:1e15. https://doi.org/ 10.3390/ani9070437.
Rommers JM, Boiti C, Brecchia G, Meijerhof R, Noordhuizen JPTM, Decuypere E, et al. Metabolic adaptation and hormonal regulation in young rabbit does during long-term caloric restriction and subsequent compensatory growth. Anim Sci 2004;79:255e64. https://doi.org/10.1017/S1357729800090111.
Rosell JM, de la Fuente LF. Culling and mortality in breeding rabbits. Prev Vet Med 2009;88:120e7. https://doi.org/10.1016/j.prevetmed.2008.08.003.
Saini RK, Keum YS. Omega-3 and omega-6 polyunsaturated fatty acids: dietary sources, metabolism, and significance d a review. Life Sci 2018;203:255e67. https://doi.org/10.1016/j.lfs.2018.04.049.
Samadian F, Towhidi A, Rezayazdi K, Bahreini M. Effects of dietary n-3 fatty acids on characteristics and lipid composition of ovine sperm. Animal 2010;4:2017e22. https://doi.org/10.1017/S1751731110001308.
Swierstra EE, Foote RH. Duration of spermatogenesis and spermatozoan transport in the rabbit based on cytological changes, DNA synthesis and labeling with tritiated thymidine. Am J Anat 1965;116:401e11. https://doi.org/10.1002/ aja.1001160206.
Tariq MR, Khan MI, Ahmad Z, Ahmed S, Sameen A, Javed MS. Development of healthier rabbit meat by supplementation of linseed in the feed and its impact on human blood lipid pro fi le. J Food Process Preserv 2017:e13194. https:// doi.org/10.1111/jfpp.13194.
Van Tran L, Malla BA, Kumar S, Tyagi AK. Polyunsaturated fatty acids in male ruminant reproduction – a review. Asian-Australas J Anim Sci 2017;30:622e37. https://doi.org/10.5713/ajas.15.1034.
Trebu�sak T, Levart A, Volj�c M, Toma�zin U, Pirman T. The effect of linseed oil sup-
plementation on performance, fatty acid composition and oxidative status of rabbits. Acta Agric Slov 2011;98:119e25. https://doi.org/10.2478/v10014-011- 0028-2.
Trebu�sak T, Fazarinc MV, Salobir J, Pirman T. The effect of substitusion of palm fat
with linseed oil in the lipid peroxidation, antioxidative capacity and intestinal morphology in rabbits (Oryctolagus cuniculus). Animals 2019;9:830. https:// doi.org/10.3390/ani9100830.
Wu L. Rabbit meat trade of major countries: regional pattern and driving forces.
World Rabbit Sci 2022;30:69e82. https://doi.org/10.4995/WRS.2022.13390.
Zerani M, Boiti C, Dall’Aglio C, Pascucci L, Maranesi M, Brecchia G, et al. Leptin receptor expression and in vitro leptin actions on prostaglandin release and nitric oxide synthase activity in the rabbit oviduct. J Endocrinol 2005;185: 319e25. https://doi.org/10.1677/joe.1.05983.