

Mattioli S.1*, Barbato O.2, Ricci G.2, Angelucci E.1, Di Federico F.1, Brecchia G.3, Castellini C.1, Collodel G.4, Dal Bosco A.1
1Dept. of Agricultural, Environmental and Food Science, University of Perugia, Borgo XX Giugno 74,
06125, Perugia, Italy
2Dept. of Veterinary Medicine, University of Perugia, Via San Costanzo 4, 06126 Perugia, Italy
3Dept. of Veterinary Medicine and Animal Sciences, University of Milan, Via dell’Università 6, 26900 Lodi,
Italy
4Dept. of Molecular and Developmental Medicine, University of Siena, 53100 Siena, Italy
*Corresponding author: simona.mattioli@unipg.it
ABSTRACT
The Omega Rabbit project (Ω RABBIT) aims to develop a new quality food product “Ωrabbit meat” assessing the impact of different n-3 fatty acids source (flax derived products or algae as Padina pavonica) on rabbit metabolism and in turn on meat quality. Moreover, the research intended to evaluate also the effect of 5% linseed and 5% linseed + 0.2% algae dietary addition on the steroid hormone profile (hormone concentration and receptors) of rabbit bucks. Forty-five New Zealand White male rabbits (6 mo) were divided into 3 dietary groups (15 animals/group): control (CNT) fed with a commercial diet, linseed group (L5%) and linseed-algae group (L5%PP) fed with the commercial diet supplemented with 5% linseed alone and 5% of linseed plus 0.2% of freeze-dried Padina pavonica algae, respectively, for 110 days (50 adaptation + 60 experimental). Semen and Blood was collected every week. Semen and blood testosterone was assessed by radioimmunoassay kits, whereas α and β-estrogens receptors (ER) by immune-fluoresce assay. The supplemented diets have an impact in the ER concentrations: L5% and L5%PP groups showed also a higher expression of sperm β ER than CNT. The diets affect also the hormonal profile of rabbit bucks: testosterone plasma concentrations were higher at 14 days (experimental period) of dietary administration in L5%PP group, whereas at 21 days the highest one was recorded for L5%. Low values respect to the CNT were recorded with linseed administration after 21 days, mainly due to the modulation of estrogen receptors. Furthermore, the reduction of testosterone due to linseed administration suggested a competition in steroids hormone. The 17-β etradiol evaluation will also be analyzed to better explain such competitive effects.
Key words: rabbit, linseed, algae, metabolism, estrogens.
INTRODUCTION
The Omega Rabbit project (Ω RABBIT) aims to develop a new quality food product “Ωrabbit meat” assessed the impact of different n-3 fatty acids source (flax derived products or algae as Padina pavonica) on rabbit metabolism and in turn on meat quality. It is well documented that linseed supplementation affects rabbit estrogen profile due to the phytoestrogens administration (Mattioli et al., 2017). Sex steroids are key reproductive system hormones in both sexes. Estrogens have always been considered the female sex steroid hormones and androgens as their male counterparts (Dewaele et al., 2022). However, it has been clear for many decades that the situation is more complex; on the one hand because the synthesis of estrogens is made from androgens, thus implying the presence, at least transitory, of androgens in females, and on the other hand because estrogens are also produced by the testes of mammals where their roles remain to be elucidated. This research aimed to evaluate the effect of 5% linseed and 5% linseed + 0.2% algae dietary addition on the testosterone plasmaconcentrations and α and β-estrogens receptors (ER) expression of rabbit bucks.
MATERIALS AND METHODS
Animals and experimental design
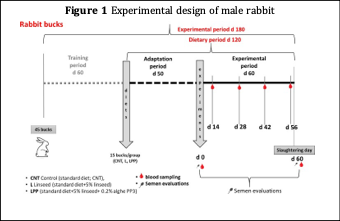
At a rabbit farm affiliated to Perugia University (Az. Agricola Brachino Patrizia), 45 male rabbits of the New Zealand White breed (6 months old) were housed in single cages and divided into 3 dietary groups (15 animals/group): the control group (CNT), was fed with a standard (commercial) diet, the linseed group (L5%) was fed with a standard diet enriched with 5% linseed and the linseed-algae group (L5%PP) was fed with a diet supplemented with freeze-dried 0.2% Padina pavonica algae + 5% linseed (Figure 1).
The diets were formulated to be isoenergetic and were supplied by a feed industry for experimental purposes (Martini srl). Ingredients and chemical composition are reported in Table 1. The diets were administered for a total of 110 days (50 adaptation + 60 experimental; Figure 1). Semen and blood was collected every week starting from T1 (7 days of the experimental period) up to T8 (48 days of experimental period). Blood samples were taken from the ear vein in 5 mL EDTA tubes. For the testosterone determination, plasma was separated by centrifugation at 2700 x g for 10 min and stored at -20° C until the next assay. Semen was taken used an artificial vagina as reported by Boiti et al. (2005).
Chemical analyses
Total testosterone was measured in duplicate by commercially available radioimmunoassay kits (Beckman Coulter – IFU-IM1119-01). The intra- and inter-assay coefficients of variation (CVs) were 10.4% and 17.8%, respectively. The lower limit of detection was 0.04 ng/mL.
The sperm of rabbit bucks fed the experimental diets were treated with 10% buffered formalin for 24 h at 4 °C and then washed in water for 1 h ad detailed in Castellini et al. (2022). Specimens were treated overnight at 4 °C with the primary antibodies anti-α or β- estrogens receptors diluted 1 : 20. After three washes for 10 min in phosphate-buffered saline (PBS), the slides (excluding those treated with conjugated primary antibody) were incubated with goat anti-rabbit antibody Alexa Fluor® 488 conjugate (Invitrogen, Thermo Fisher Scientific, Carlsbad, CA, USA), diluted at 1 : 100, for 1 h at room temperature. The slides were washed three times with PBS and treated with 4′,6-diamidino-2-phenylindole (DAPI, Sigma-Aldrich, Milan, Italy) for 10 min, followed by washing with PBS for 10 min. Finally, the slides were mounted with 1,4-diazabicyclo[2.2.2]octane (DABCO, Sigma-Aldrich, Milan, Italy).
Statistical Analysis
The statistical analysis of All the numerical results was carried out with the ANOVA model where the fixed effects evaluated were time, diet and their interaction. For the multiple comparison, the Bonferroni ad hoc test was used, with an alpha value of 5%.
Table 1. Ingredients (%) of the experimental diets for the rabbit bucks
| CNT | L5% | L5%PP | |
| Ingredients | |||
| Bran Wheat | 25.07 | 24.87 | 24.92 |
| Barley | 13.33 | 13 | 13 |
| Sunflower seed meal | 12 | 11.67 | 11.5 |
| Alfalfa | 10.83 | 13 | 13 |
| Sunflower husks | 10 | 10 | 10 |
| Beet pulp | 7.5 | 5.67 | 5.5 |
| Extruded linseed | – | 5 | 5 |
| Full-fat soybean | 5 | 2.95 | 3.1 |
| Wheat straw | 4.17 | 2 | 2 |
| Molasses cane | 3 | 3 | 3 |
| Wheat | 2.5 | 2.5 | 2.5 |
| Grape pips meal | 2.33 | 1.67 | 1.67 |
| Soya hulls | 0 | 1.67 | 1.67 |
| Calcium carbonate | 1.6 | 1.48 | 1.42 |
| Soybean oil | 0.78 | – | – |
| Sodium chloride | 0.4 | 0.4 | 0.4 |
| Palm oil | 0.33 | – | – |
| Carboxymethylcellulose | 0.3 | 0.3 | 0.3 |
| Oligo-vitamin supplement1 | 0.25 | 0.25 | 0.25 |
| Alga PP | – | – | 0.2 |
| Lysine HCl | 0.16 | 0.17 | 0.17 |
| Liquid acidifier2 | 0.15 | 0.15 | 0.15 |
| Magnesium oxide | 0.1 | 0.1 | 0.1 |
| Methionine hydroxyanalog | 0.06 | 0.07 | 0.07 |
| Liquid choline | 0.05 | 0.05 | 0.05 |
| Vitamin E 50% | – | 0.03 | 0.03 |
| L Threonine | 0.03 | 0.01 | 0.01 |
| DL Methionine | 0.03 | – | – |
| Chemical composition | |||
| Protein | 16.5 | 16.5 | 16.5 |
| Lipids | 3.62 | 3.93 | 3.93 |
| Fiber | 17.16 | 16.79 | 16.79 |
| Ash | 7.99 | 8.09 | 8.09 |
| ED rabbit (kcal) | 2350.00 | 2350.00 | 2350.00 |
1Vitamin Mineral premix composition: Vitamine A, D3, E, K3, B1, B2, B6, B12, Biotin, Niacinamide, Folic acid, Calcium pantothenate.
2Liquid acidifier composition: Formic acid 75%
RESULTS AND DISCUSSION
Effect of experimental diets on semen estrogen raptors distribution
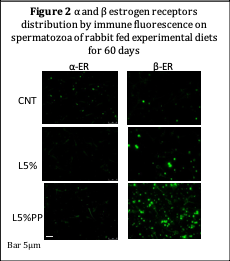
Sperm of L5% and L5%PP groups showed higher expression of β ER than CNT but not for the α ER (Figure 2). Furthermore, a different distribution on sperm surface was also recorded: β ER were mainly located in the head, while α ER in the midpiece of sperm, probably due to their different functions in sperm.
Effect of experimental diets on testosterone concentration
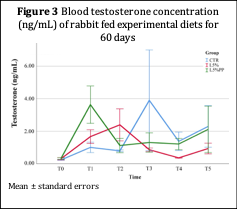
Blood testosterone (Figure 3) showed a higher concentration at 14 days of dietary administration in L5%PP group, whereas at 21 days the highest one was recorded for L5%.
Low values respect to the CNT were recorded with linseed administration after 21 days.
CONCLUSIONS
Dietary administration of 5% flaxseed widely affected the hormone profile of rabbit bucks, mainly due to the modulation of estrogen receptors. Furthermore, the reduction of testosterone due to linseed administration suggested a competition in steroids hormone. The 17-β estradiol evaluation will also be analysed to better explain such competitive effects.
ACKNOWLEDGEMENTS
Research was funded by PRIMA ΩRABBIT project.
REFERENCES
Boiti, C., Castellini, C., Besenfelder, U., Theau-Clément, M., Liguori, L., Renieri, T., & Pizzi, F. 2005. Guidelines for handling of rabbit bucks and semen. World Rabbit Science, 13(2), 71-91.
Castellini, C., Mattioli, S., Moretti, E., Cotozzolo, E., Perini, F., Dal Bosco, A., … & Collodel, G. 2022. Expression of genes and localization of enzymes involved in polyunsaturated fatty acid synthesis in rabbit testis and epididymis. Sci. Reports, 12(1), 2637
Dewaele, A., Dujardin, E., André, M., Albina, A., Jammes, H., Giton, F., … & Pannetier, M. 2022. Absence of Testicular Estrogen Leads to Defects in Spermatogenesis and Increased Semen Abnormalities in Male Rabbits. Genes, 13(11), 2070.
Mattioli, S., Ruggeri, S., Sebastiani, B., Brecchia, G., Dal Bosco, A., Mancinelli, A. C., & Castellini, C. 2017. Performance and egg quality of laying hens fed flaxseed: Highlights on n-3 fatty acids, cholesterol, lignans and isoflavones. Animal, 11(4), 705-712.